
6月9日,我院王应祥课题组联合复旦大学客座教授北卡罗来纳大学教堂山分校Gregory Copenhaver研究组在PLOS Genetics杂志在线发表题为“The cohesin loader SCC2 contains a PHD finger that is required for meiosis in land plants”的研究论文,揭示了植物减数分裂过程中SCC2参与招募黏连蛋白的新机制。
减数分裂是真核生物有性生殖的必需环节,其过程需要一类黏连蛋白(Cohesin)捆绑姐妹染色单体,以确保后续同源染色体之间配对、联会、重组和分离的有序进行。黏连蛋白是真核生物中高度保守的多亚基蛋白复合物,有丝分裂中主要负责捆绑DNA复制后形成的姐妹染色单体,维持基因组的稳定及染色体的正确分离。酵母有丝分裂的研究发现,SCC2/SCC4复合物(Sister Chromatid Cohesion 2/4)负责染色体上Cohesin的装载。其中,SCC2促进Cohesin装载到染色体上,而SCC4决定了染色体上的结合位点。但由于Cohesin及SCC2/SCC4复合物功能的重要性,动植物中SCC2/SCC4完全缺失会胚胎致死,限制了对生物学功能的深入研究。目前对于植物减数分裂中SCC2/SCC4如何帮助招募Cohesin的机制尚不清楚。
此研究利用Copenhaver教授团队化学诱变构建的拟南芥突变体库,从中筛选到一个减数分裂异常突变体。通过基因组重测序发现,其SCC2基因的第22外显子3’端发生单碱基突变,导致转录本的错误剪切及蛋白翻译的提前终止。细胞学分析该突变体的减数分裂过程,发现同源染色体联会、重组和分离均不正常;进一步分析发现减数分裂重组的起始正常,但是减数分裂特异的Cohesin亚基SYN1的装载异常。遗传分析结果显示SCC2与多个Cohesin相关基因SYN1、WAPL1/2处于同一通路,并参与RAD51介导的以姐妹染色单体为模板的重组修复。
该研究进一步证明SCC2的N端与SCC4相互作用,暗示植物减数分裂过程中SCC2-SCC4可能也通过形成复合体来发挥功能。然而,通过特异降低减数分裂细胞中SCC4的表达量,虽然转基因植株的育性明显降低,但未发现减数分裂异常,表明植物减数分裂可能并不需要SCC4,同时揭示了SCC4在植物有丝分裂和减数分裂过程中的功能分化。
此外,不同于动物SCC2蛋白,陆生植物SCC2蛋白具有一个特有且保守的PHD(plant homeodomain)结构域,体外生化实验证明SCC2的PHD结构域能够结合未修饰及甲基化的组蛋白H3、H4和H2A。体内功能实验证明,AtSCC2的PHD结构域是正常减数分裂所必需,而有丝分裂中则不需要该结构域。
综上所述,该研究提出了SCC2参与减数分裂黏连蛋白复合体招募的工作模型(见下图)。在动植物的减数分裂间期(Pre-meiosis),SCC2与SCC4共同参与招募Cohesin到染色体上,其中SCC2通过C端的“钩子”帮助Cohesin的装载,SCC4负责决定Cohesin在染色体上的结合位点。进入减数分裂后,SCC2-SCC4在减数分裂特异的Cohesin装载或替换中出现了功能分化。动物SCC2-SCC4可能仍然以复合体执行减数分裂特异的Cohesin替换,其位点特异性可能由SCC4决定;而植物SCC2通过不依赖于SCC4的方式,利用特异PHD结构域结合组蛋白,从而决定Cohesin的替换位点。该研究首次揭示了植物减数分裂中SCC2招募黏连蛋白的作用机制,为动植物减数分裂过程中SCC2介导黏连蛋白招募的异同提供了依据。
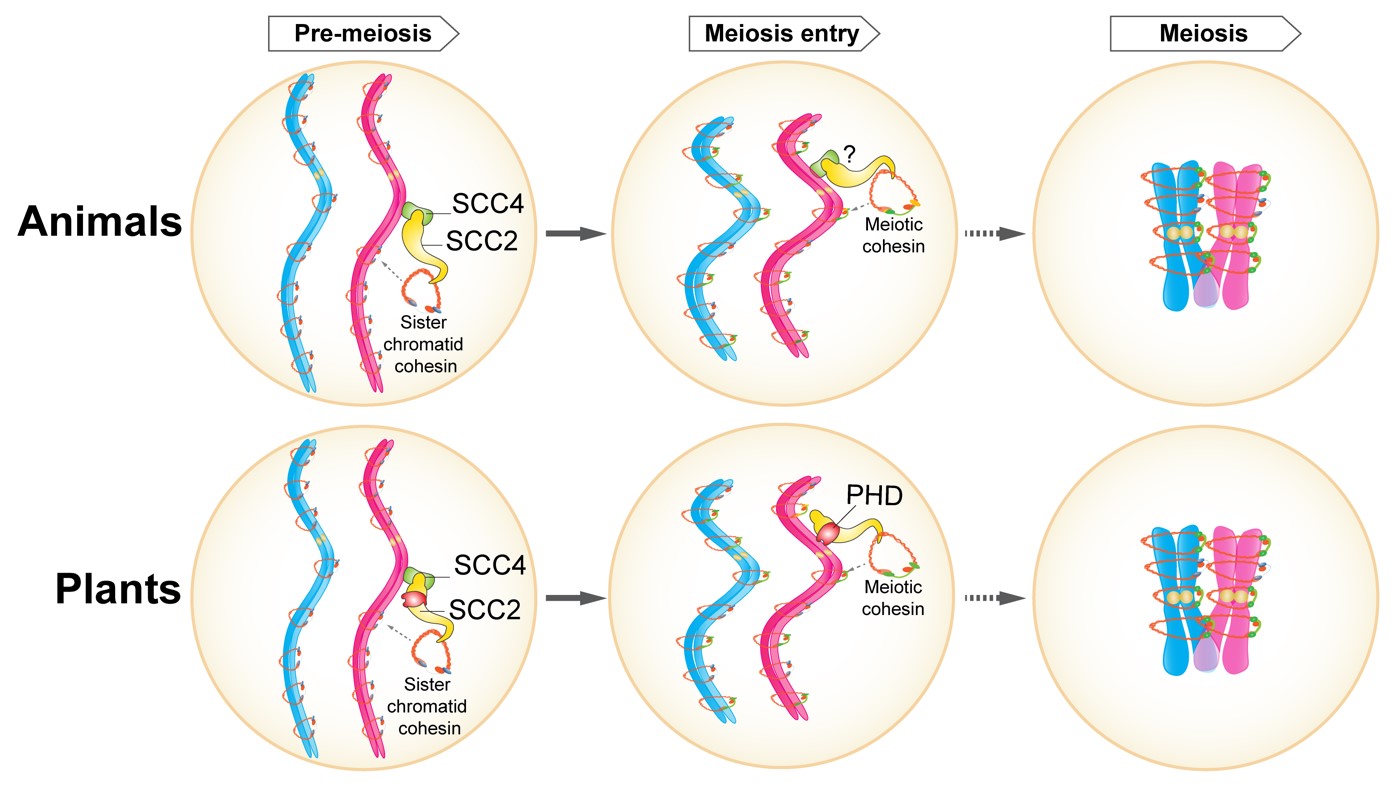
图注:动植物减数分裂中SCC2装载黏连蛋白复合体的作用模型
我院15级博士生王宏宽及17级博士生徐婉约为此文的并列第一作者,遗传工程国家重点实验室、遗传与发育协同创新中心,生物多样性和生态工程教育部重点实验室的王应祥研究员与美国北卡罗来纳大学教堂山分校的Gregory Copenhaver教授为文章的共同通讯作者,美国宾州州立大学的马红教授参与并对该课题提供了大量帮助。该研究获得了国家自然科学基金委员会和复旦大学遗传工程国家重点实验室,及美国国家自然科学基因项目的共同资助。
全文链接:https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1008849