
王应祥课题组在减数分裂交换分布模式的调控机制研究取得重要进展
2021年11月18日,国际学术期刊PNAS在线发表了生命科学学院王应祥研究组题为“Regulation of interference-sensitive crossover distribution ensures crossover assurance in Arabidopsis”的研究论文(https://www.pnas.org/content/118/47/e2107543118)。该研究发现减数分裂交换在染色体间的分布模式的分子遗传机制,同时揭示了减数分裂交换保障现象的调控机制。

真核生物通过减数分裂产生染色体数目减半的配子(精子或卵子),精卵受精结合后,染色体数目又恢复到亲本数目,维持了基因组的稳定性。不同于有丝分裂,减数分裂涉及到同源染色体间的配对、联会和重组。重组使得同源染色体间发生交换,其正确与否直接影响染色体的分离,最终导致人类流产和染色体综合征疾病,如唐氏综合征等,以及作物的不育和产量降低。除了确保同源染色体的正确分离,交换还促使同源染色体间遗传物质发生重新组合,增加了后代的遗传多样性,为自然选择过程提供了丰富的变异基础,促进了物种的进化。此外,同源染色体交换也是遗传育种的基础,育种家们利用同源染色体交换来组合优良变异培育优良品种。已有研究表明,减数分裂同源染色体间交换的数目和分布模式受到严格的调控。例如模式植物拟南芥每次减数分裂形成约10个交换,假如这10个交换在拟南芥5对同源染色体间随机分布,将会造成约10%的染色体缺乏交换,进而导致异常分离。实际上,拟南芥减数分裂同源染色体间至少能形成一个交换,否定了交换在染色体间的随机分布,这种现象被称为交换保障。交换保障现象在不同物种间高度保守,最早是科学家Darlington和Dark由1932年研究蝗虫的减数分裂染色体行为时发现的。1949年科学家Owen在研究蚊子的减数分裂过程中,正式提出了交换保障概念。虽然交换保障现象被发现至今已经将近百年,人们对于其潜在的分子调控机制仍然知之甚少。
在探索维持交换保障的分子机制的过程中,研究者首先发现拟南芥减数分裂过程,每对同源染色体至少形成一个I型交换,I型交换在染色体间并不是随机分布。而II型交换在染色体间的分布则是随机的。说明正常情形下,只需要I型交换就能维持交换保障,交换保障的缺失必然伴随着I型交换数目或者分布模式的改变。基于这种思考,同时也为了简化研究思路,研究者首先在已经发表的减数分裂相关突变体中检索I型交换数目不变,同时伴随有染色体单体产生的突变体。研究者首先锚定到了促进II型交换形成的FANCD2蛋白,fancd2突变体的减数分裂中,I型交换数目不变,但13%染色体上缺乏交换。对I型交换在染色体间的分布模式研究发现,fancd2突变体中5%同源染色体上缺乏I型交换,I型交换在染色体间的分布模式处于野生型和泊松分布的平均状态,也就是一种“半随机”状态,说明FANCD2具有促进I型交换在染色体间的正确分布维持交换保障的功能。
接着研究发者抑制II型交换数目的FANCM,FIGL1以及RMI1也具有类似于FANCD2的调控I型交换分布保正交换保障的功能,单独或同时破坏这些因子都使得I型交换的分布模式处于“半随机”状态,伴随着5%的同源染色体上缺乏I型交换(图1C)。在fancm、figl1和rmi1突变体中,虽然II型交换数目升高,但是II交换在染色体间随机分布,并不能完全保证缺乏I型交换的染色体获得交换,因此这些突变体中仍能观察到未发生交换的染色体单体。
进一步遗传分析揭示,FANCM、FIGL1、RMI1和FANCD2在调控I型交换分布,维持交换保障过程利用类似的机制。而在调控交换数目时则通过不同的途径,有意思的是FANCD2在fancm突变体背景下仍然促进II型交换形成的功能,而在figl1或rmi1突变体背景下,FANCD2则抑制II型交换的形成(图1B)。
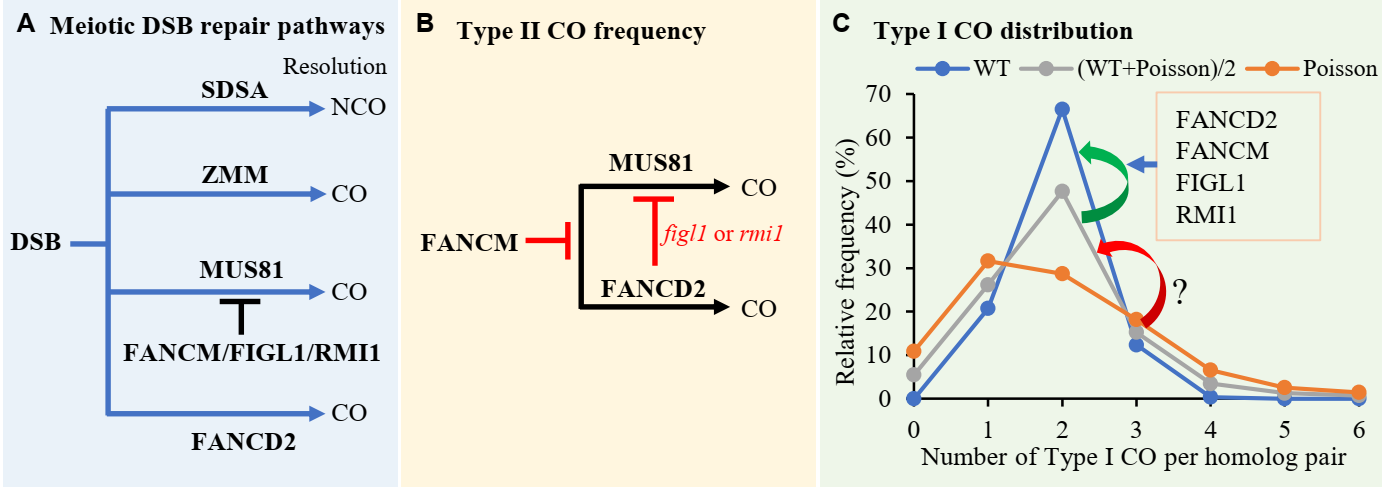
图1:拟南芥减数分裂交换数目和频率的调控模型。
综上所述,本研究以拟南芥为研究对象,揭示了多个II型交换数目调控因子具有调控I型交换在染色体间分布,维持交换保障的功能,为保障现象的分子维持机制提供了新视角,也为操纵交换分布模式和交换频率,加速作物育种提供了理论依据。
复旦大学生命科学学院王应祥课题组已毕业博士研究生李翔为文章第一作者(现工作单位为河南农业大学园艺学院),王应祥为文章通讯作者。复旦大学兼职教授、北卡罗来纳大学教堂山分校Gregory Copenhaver教授和黄霁月博士后也参与了该项工作。该研究得到了国家自然科学杰出青年基金,美国国家科学基金以及复旦大学的资助。