
mTOR信号通路是维持细胞正常生长、发育的重要通路之一,并且该通路调控异常与乳腺癌、肺癌、肝癌和结肠癌等多种肿瘤的发生发展关系密切。在细胞内的mTOR主要以mTORC1和mTORC2这两种蛋白复合体的形式存在。mLST8 是mTORC1复合体和mTORC2复合物中的共有组成成分。 mLST8不是mTORC1复合体活性激活的必需成分,而是mTORC2复合体活性激活的必需组分。但目前对于mLST8参与mTORC2 复合体的组装和动态调控的了解还不够透彻。
Keap1是Cullin 3 家族 E3 泛素连接酶的一个底物识别亚基,Cullin 3 泛素 E3 连接酶复合体(Cul3-RBX1-BTB)是由支架蛋白 Cullin 3,催化亚基 RBX1 和一个含有 BTB 结构域的底物识别亚基组成。 Keap1在肾癌、胃癌、结直肠癌、肝细胞癌和肺癌等多种肿瘤均发生不同频率的失活突变。 Keap1是一个研究得较为深入的肿瘤抑制基因,已经报道 Keap1的生物学功能大部分集中在其对底物蛋白氧化应激相关转录因子—NRF2的泛素化降解,进而调控细胞的氧化应激反应。但目前对于 Keap1介导的非NRF2相关的生物学过程的研究仍然不够深入。
2023年9月6日,复旦大学生命科学学院王陈继团队、同济大学附属第一妇婴保健院高昆团队和内蒙古自治区人民医院莫仁团队合作在Redox Biology在线发表了题为Disruption of the Keap1-mTORC2 axis by cancer-derived Keap1/mLST8 mutations leads to oncogenic mTORC2-AKT activation的研究论文(图一)。该研究发现CRL3Keap1 E3泛素连接酶通过一个保守的ETGE基序与mLST8结合, 介导了mLST8的非降解型泛素化修饰,从而抑制mTORC2复合物的组装以及mTORC2-AKT活性的分子机制。

鉴于mLST8是mTORC2复合体的关键成分,研究人员通过亲和纯化的方法分离mLST8蛋白复合体,然后用质谱分析的方法鉴定新的mLST8结合蛋白,进而推断新的mTORC2调控模式。通过该方法,研究人员首次发现并确证了Keap1是一个新的mLST8互作蛋白。Keap1通过识别mLST8蛋白序列中的一个典型Keap1 结合基序(300-ETGE-303 aa)与mLST8相互作用,并且可以对mLST8进行泛素化修饰,但是与NRF2的泛素化降解不同,此修饰不影响mLST8的蛋白水平。在肿瘤中,mLST8的ETGE基序内发生的氨基酸突变(G302R和E303D)减弱了 mLST8和Keap1 的相互作用,以及 Keap1 对 mLST8的泛素化修饰。另外,癌症来源的 Keap1 突变体和 mLST8 的互作显著降低,并且Keap1 对 mLST8 的泛素化修饰也明显减弱。 肿瘤中Keap1失活突变、或mLST8的ETGE基序发生突变以及细胞处于氧化应激的条件下,可以解除Keap1对mTORC2-AKT信号通路的抑制作用,
后续的实验证实,细胞在正常状态下,Keap1 介导 mLST8 的非降解型泛素化修饰,导致mLST8 脱离 mTORC2 复合体,mTORC2 复合体组装受阻,活性下降,其下游AKT 激酶活性保持在较低水平;在某些肿瘤中以及细胞在氧化应激的条件下,Keap1 或者mLST8失活, mLST8 不能被 Keap1 泛素化修饰,游离的 mLST8 更多地参与 mTORC2 复合体组装,mTORC2 复合体及其下游的 AKT 激酶活性上升,进而促进细胞的增殖、迁移、侵袭、锚定非依赖性生长、移植瘤体生长和细胞糖代谢重构等肿瘤恶性转化表型(图二)。
总之,这项工作本研究首次提出了“Keap1-mTORC2-AKT”信号调节轴失调在肿瘤发生发展过程中的重要作用,为mTOR、AKT抑制剂应用于Keap1突变型肿瘤的靶向治疗提供了理论基础。
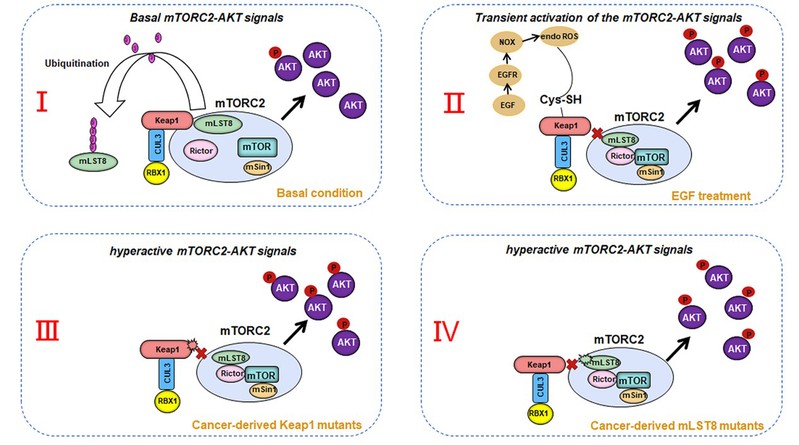
图二、Keap1-mTORC2-AKT 信号通路的动态调控分子机制
复旦大学生命科学学院博士生陈莹佶、焦冬月,硕士生何慧莹为该论文共同第一作者。王陈继、高昆和莫仁为该论文的共同通讯作者。
原文连接:https://doi.org/10.1016/j.redox.2023.102872